

Feb 2004, Sep 2014
Part 3
5. Anatomy and origins
6. Theosophy: fallen angels,
fallen apes
A comparison of the skeletal and muscular features of living apes and humans shows that apes have developed a more complex and specialized anatomy, while humans have preserved a primitive mammalian simplicity, with only the cerebrospinal system, necessary for the manifestation of self-conscious intelligence, being highly developed. If apes and humans descended from a common ancestor, that ancestor must have had a more generalized anatomical structure than modern apes. It is widely recognized that the hypothetical common ancestor of humans and chimpanzees must have been more humanlike than chimplike.1 Fossil apes predating this ancestor also tend to display simpler, more hominin-like features, while still possessing certain ape specializations to varying degrees.
Ramapithecus is a fossil primate about 1.2 m (4 ft) tall, dating from the middle and late Miocene (about 16.6 to 5.3 million years ago). For a time in the 1960s and 70s it was thought to be the first direct ancestor of modern humans, as its jaw and dentition seemed to be transitional between those of apes and humans (i.e. more humanlike than those of modern apes). The age of the first fossils found fitted well with the then-prevailing notion that the ape-human split had occurred at least 15 million years ago. However, the final split is now believed to have taken place only 6 to 8 million years ago. Following further fossil finds, including a complete jaw, the theory that Ramapithecus was a human ancestor was largely abandoned by the early 1980s. Its fossils were found to resemble those of the fossil primate genus Sivapithecus, which is now regarded as ancestral to the orangutan.2
Fossils of the primate Oreopithecus, which inhabited swampy regions, are found in late Miocene deposits in East Africa and early Pliocene deposits in southern Europe (11.6 to 3.6 million years ago). A complete skeleton of this 1.2-m-tall creature was discovered in 1958 and was found to have a number of curiously humanlike traits in its teeth, jaws, skull and hipbone. Its relation to other primates has been a matter of some debate and confusion, as it combines primitive and advanced features, which seem to ally it with the Old World monkeys on the one hand and with the advanced, humanlike apes on the other. It is no longer seen as belonging to the human line of descent, but is widely regarded as a specialized side-branch of primate evolution. It had long arms adapted for suspensory behaviour. The extent to which it was capable of bipedal locomotion is a matter of controversy.3 Gigantopithecus, too, displays several humanlike traits, including reduced canine teeth, and as noted in section 4, a few scientists have concluded that it was bipedal.
The tree-living, fruit-eating dryopithecine apes from the Miocene are widely believed to be close to the origin of the hominid line of descent. But even the earliest dryopithecines have the specialized dentition characteristic of apes, and some scientists think it unlikely that such a specialization could be lost by a reversal to the primitive condition.4 Like modern humans, the dryopithecines did however have small brow ridges. Yet Australopithecus, Homo erectus and the Neanderthals, along with the modern apes, have large brow ridges. Louis Leakey thought it unlikely that Miocene apes with no brow ridges could give rise to early hominids with heavy brow ridges, who in turn gave rise to modern humans with virtually nonexistent brow ridges.
Furthermore, in H. sapiens the brow ridge is made up of two components, while in Australopithecus, H. erectus and the Neanderthals the large brow ridges are most often composed of a single barlike mass of bone running horizontally over the eye sockets. To Louis Leakey, the presence of such barlike brow ridges ‘suggested not an ancestral stage in human evolution but a side-branch that has become more specialized, in this respect, than any Homo sapiens type’. In addition, Miocene apes tend to have thin skulls, while the australopithecines, H. erectus and the Neanderthals have relatively thick skulls. Modern humans have thin skulls, which would require another evolutionary reversal. Leakey felt that the specializations displayed by the australopithecines and H. erectus ruled them out as ancestors leading to modern humans.5 As indicated in section 1, Tattersall and Schwartz take a similar view.
A trend towards lighter and more delicate skeletons is known as ‘gracilization’. Scientists, like Leakey, who see gracilization as implausible have responded by placing H. erectus and the Neanderthals, and in some cases the australopithecines, on different branches of the evolutionary tree to that of modern humans. However, other Darwinists do not see evolutionary reversals as particularly anomalous. The idea that random genetic mutations (even after harmful ones have been eliminated by natural selection) will produce progressive evolutionary change is improbable enough, and the idea that a particular sequence of mutations will later be repeated but in reverse order is even more improbable, but Darwinists simply reply that improbable does not mean impossible. Some scientists have linked gracilization to neoteny, the biological condition in which juvenile anatomical traits are retained in adults (see below). But giving a problem a newfangled name is not the same as explaining it.
Björn Kurtén was another anthropologist who believed that more specialized anatomical characteristics must develop from less specialized features. In Not from the Apes (1972), he argued that the contrasts between apes and humans in anatomy were so great that the two lineages must have diverged over 35 million years ago. He held that the earliest ancestor of modern man was undoubtedly apelike and tree-living but was never what could properly be called an ape. He thought that the cat-sized Oligocene primate Propliopithecus, with its striking humanlike traits, might be ancestral to Ramapithecus – which at the time he was writing was widely considered to be a hominid (or ‘hominin’ in modern terminology) – and to all later hominids, including the australopithecines, while other apelike primates from the Oligocene gave rise to Dryopithecus and to the later monkeys and apes. He concluded that ‘the most logical answer suggested by the fossil evidence is this: hominids are not descended from apes, but apes may be descended from hominids’. By this he meant that the living apes evolved from early ancestors of man, i.e. apelike hominids, who in turn evolved from an earlier primitive apelike primate.6
In an article written in 1981, John Gribbin and Jeremy Cherfas asked: ‘What if, instead of man being descended from the apes, the apes are descended from man?’7 However, as in the case of Kurtén’s book, their theory turns out to be far less radical than the title might suggest. They accept that ‘man’ (Homo) descended from a branch of the australopithecines, but argue that instead of becoming extinct a million years ago, the remaining australopithecines evolved into the modern chimpanzee and gorilla, whose ancestors are currently unknown. This means that our supposed apelike ancestors evolved into bipedal hominids (Australopithecus), some of whom later reverted to life in the trees. Gribbin and Cherfas comment: ‘Whatever the small genetic changes needed to accomplish the anatomical reshufflings that produced an upright ape, they could surely have been equally easily reversed.’ They admit that their proposal ‘may seem like a wild flight of fancy, the over-vivid imaginings of two armchair anthropologists’.
There is a widespread tendency to overemphasize the bodily resemblances between humans and apes while playing down the significant host of dissimilarities. In the early 20th century, anatomist Frederick Wood Jones cited various anatomical facts as evidence that humans could not have evolved from the apes or their direct ancestors. He argued that it was difficult to imagine that in a whole series of uncorrelated features humans could have passed from a primitive condition to the specialized pithecoid condition, and subsequently have reverted to the primitive condition. He believed that man originated in the early Tertiary from a small arboreal creature, closely resembling the monkeylike tarsier of today, and that the australopithecines are not part of our ancestry.
The evidence pointing to the primitiveness of the human stock is summarized by G. de Purucker in his book Man in Evolution, and includes the following:8
Neoteny (literally ‘holding youth’) means the retention of the juvenile features of an ancestral species in the adult form of a descendant species. It results from a slowdown in the rate of physical maturation, so that the individual is frozen in a physically juvenile state upon reaching sexual maturity. As noted above, humans have several physical features that seem to be more juvenile or primitive than the same features in other primates. Many external characteristics of apes differ from those of humans, but these differences tend to become less pronounced as we retrace the bodily development of apes backward to the juvenile and embryonic stages. This suggests that, as far as anatomy is concerned, apes are more specialized and further evolved than humans, and that many human features are ‘retarded’ in their development. In the 1920s anatomist Louis Bolk went so far as to proclaim that humans are actually reproductively mature fetuses, or upright-walking embryos – a theory he called ‘fetalization’.
Fig. 5.1. A young chimpanzee has a very humanlike face, but adults develop a prominent muzzle.1
In 1866 arch-Darwinist Ernst Haeckel formulated the ‘biogenetic law’, which states that the development of the embryo (ontogeny) recapitulates the evolutionary ancestry of the stock in question (phylogeny). He argued that as an organism passes through embryonic development it retraces every adult stage of its evolutionary ancestry, because an organism evolves by tacking on new stages to its process of embryonic development. Biologists soon discarded this idea, recognizing that embryology is not a strict replay of ancestry as evolution can affect all phases of development, removing developmental steps as well as adding them. Nevertheless, it is still accepted that many of the stages that embryos pass through can be understood as remnants of their evolutionary past. Theosophy agrees with this,2 though it rejects the neo-Darwinian theory that new stocks of creatures emerge through the continuous transformation of one physical form into another as a result of random mutations and natural selection.
The Darwinian interpretation of neoteny is that humans evolved by retaining the youthful (more humanlike) features of our supposedly apelike ancestors. From a theosophical perspective, the humanlike features of embryonic and juvenile apes are a sign of their semi-human ancestry (see next section); apes are humans that have evolved physically and devolved mentally.

Fig. 5.2. Comparison of the skulls of a human (left column) and the capuchin monkey (right column), representing most anthropoid primates. The juveniles of the human and the monkey (bottom) are similar in that both have a large rounded head, a relatively unprojecting snout, large eyes, weak brow ridges, and small jaws and teeth. The adult human typically retains these juvenile features, while the adults of monkeys and other mammals tend to develop larger and more projecting lower faces and brow ridges.3
The suture (or junction) between the premaxilla (the front part of the upper jawbone carrying the incisor teeth) and the maxillary bones fuses at an early stage in the development of the human embryo. Wood Jones argued that this precludes an origin from monkeys, apes and even from the australopithecines, in which the suture closes later and the suture lines remain visible on their faces. Human and ape teeth are similar in many ways, but differences do exist and are more pronounced in the second dentition (permanent teeth) than in the milk teeth. Juvenile characteristics are retained in humans, while the apes go on to develop large canines, thin enamels, and the dental geometry that gives them their ‘canine profile’.4
Adult monkeys and gorillas are hairy all over, but monkeys are born with hair only on their heads and backs, and gorillas with hair only on their heads. Newborn humans also have hair only on their heads, and – compared with adult monkeys and apes – adult humans are not much hairier. From a Darwinian perspective, the rate at which hair spreads to cover the body has become significantly retarded in humans. Theosophically, the rate at which hair spreads has accelerated in apes.
At a certain stage of its development the human fetus is covered with a downy layer of hair that sometimes lingers on the skin of newborn babies. Darwinists interpret this to mean that the human fetus passes through an ‘ape stage’. However, this fetal hair has nothing to do with animal hair; it is entirely human hair. In reality, the hair of animals grows onto the first layer of the human-type hair – another indication that apes have evolved beyond humans.5
Recognizing that many features of man’s anatomy were ‘retarded’ while our mental development has undergone accelerated development, E.D. Cope wrote:
[I]n the structure of his extremities and dentition, he agrees with the type of Mammalia prevalent during the Eocene period. Hence in these respects he resembles the immature stages of those mammals which have undergone special modifications of limbs and extremities ... [I]n the shape of his head man resembles the embryos of all Vertebrata, in the protuberant forehead, and vertical face and jaws. In this part of the structure most Vertebrata have grown further from the embryonic type than has man, so that the human face may be truly said to be the result of retardation. Nevertheless, in the structure of his nervous, circulatory, and for the most part, of his reproductive system, man stands at the summit of the Vertebrata. It is in those parts of his structure that are necessary to supremacy by force of body only, that man is retarded and embryonic.6
 The theory of initial bipedalism,1 as
championed by François de Sarre of the Study and Research Centre for
Initial Bipedalism (CERBI), states that instead of being a fairly recent
evolutionary transformation of an apelike ancestor, humans are an extremely
ancient stock and have remained morphologically and anatomically closest
to the original vertebrate from which all others descend. In De Sarre’s
view, this prototype was a hypothetical tiny marine creature that lived some
60 million years ago.
The theory of initial bipedalism,1 as
championed by François de Sarre of the Study and Research Centre for
Initial Bipedalism (CERBI), states that instead of being a fairly recent
evolutionary transformation of an apelike ancestor, humans are an extremely
ancient stock and have remained morphologically and anatomically closest
to the original vertebrate from which all others descend. In De Sarre’s
view, this prototype was a hypothetical tiny marine creature that lived some
60 million years ago.
He describes the mainstream evolutionary history of man as ‘a gigantic farce, based on erroneous observations and old prejudices’, and says that since the palaeontological record is so incomplete we have to rely mainly on embryology and comparative anatomy to discover our real origins. Rather than interpreting man’s primitive (‘juvenile’) structure to mean that humans are fetal apes that have matured, De Sarre argues, as does theosophy, that apes descend from man and have evolved anatomically beyond the point where human development ceased.
Humans are the only known mammals that are habitually bipedal. Significantly, in the embryos of humans, other primates, and many other mammals, the foramen magnum (the opening in the skull through which the spinal column passes) is positioned centrally. In adult humans it retains this position – which is required for bipedalism – whereas in other animals, including apes, the opening migrates backward, as growth proceeds, to the position needed by a quadruped.2
A related fact is that in all mammal embryos, including humans and apes, the angle between the plane of the face and the plane of the base of the skull, through which the spinal cord passes, is extremely flexed – essentially 90°. This angle remains well flexed in adult humans (120°), this being connected with our bipedal mode of locomotion. In adult animals, on the other hand, the angle opens up, rising to 140° in apes, and up to 180° in fully quadrupedal mammals.
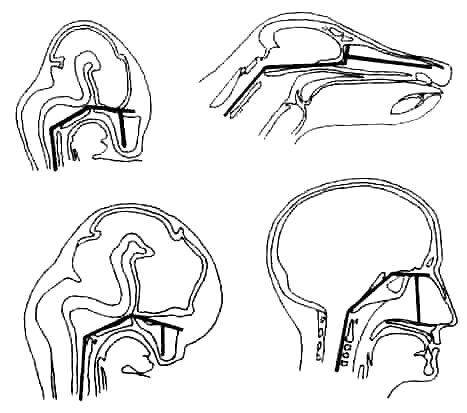
Fig. 5.3. Representation of the angle between the facial skeleton and the skull base in a dog (above) and a human (below), showing the fetal stage (left column) and the adult stage (right column).3
An analysis of man’s hands and feet shows that he has never been a quadruped. Apes and monkeys have prehensile feet (i.e. an opposable big toe) that are like lower hands and permit the grasping of branches etc. In addition, the third digit of their feet is the longest of the five. The human foot, geared to bipedalism, is quite different, and typically human from the time of its first appearance early in the embryo’s development. Significantly, in the embryo all primate feet begin as humanlike feet, implying that the foot of apes and monkeys is a human foot that has become prehensile through adaptation to life in the trees. In addition, the third peroneal muscle of the leg – one of the important muscles which aid humans to stand upright and to walk bipedally – is found in no other mammal whatsoever. It is found in the human embryo early in its development, suggesting that man must have had an upright posture perhaps from the very origin of the human stock.4
The human hand, with its five fingers, has remained very primitive, and cannot have developed from an animal’s paw. The structure of the human arm and hand has served as a starting point for various evolutionary transformations, such as the forelimbs of quadrupedal mammals, the wings of bats, and the pectoral fins of cetaceans. Whereas humans have a well-developed opposable thumb, the thumb of nonhuman primates is short, often emaciated, or entirely missing as a result of disuse. In some monkey species, specialization in tree-climbing has resulted in their thumb being reduced to a stump, while at the same time their arms have lengthened. Apes have retained hands that are anatomically very similar to man’s, but they often employ them for knuckle-walking.5
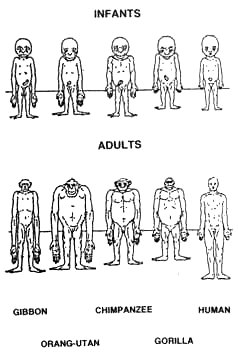
Fig. 5.4. Changes in body proportion in humans and apes. Max Westenhöfer argued that if we don’t hold any preformed opinion, this diagram rules out a simian ancestry for man.6 Apes’ forelimbs are longer than their hindlimbs as a result of specialization to life in the trees.
Palaeontologist Yvette Deloison cites the primitive structure of the human hand as evidence that the common ancestor of the australopithecines, the great apes, and humans, could not have been either arboreal or quadrupedal. The human foot, on the other hand, is specialized for bipedal locomotion. On the assumption that evolution never goes backward, she argues that our earliest primate ancestor must have had unspecialized hands and feet, and that humans and australopithecines had a common bipedal ancestor who appeared about 15 million years ago. Rather than evolving towards human bipedalism, the semi-bipedal australopithecines were therefore adapting to a new way of life in the trees.7
De Sarre adopts a similar position. Like Gribbin and Cherfas, he believes that the present anthropoid apes could be related to the australopithecines. But instead of arguing that the australopithecines were bipeds which evolved from quadrupedal ancestors but later regressed and turned into quadrupedal apes, he argues that the australopithecines had bipedal ancestors and were actually in the process of losing their bipedalism and evolving towards quadrupedalism, as part of a process of dehumanization. He regards wildmen such as Bigfoot and the Yeti as relict dehumanized hominoids.8
The theosophical teachings on evolution given out since the formation of the
Theosophical Society in 1875 are merely a general outline of the information
in the possession of the
Brotherhood of Adepts.1 This information is said
to have
been compiled and repeatedly verified by countless generations of sages
and seers, whose occult powers grant them access to the inner realms of
nature and enable them to read the records of the earth’s history
clairvoyantly. The adepts are also said to possess written records, fossils,
technological inventions and other artifacts from bygone ages. They disseminate
their knowledge in
proportion to people’s ability to respond to it wisely.
Darwinism is rooted in the materialistic assumption that the universe consists essentially of physical matter and energy, and that mind and consciousness are merely byproducts of the brain. According to the theosophic tradition, on the other hand, the physical world is the outer shell of inner worlds – astral, mental and spiritual. Likewise, every physical organism is animated by inner, subtler ‘bodies’ or souls, including an astral model-body (the template for the physical body), an instinctive and/or self-conscious mind, of widely varying degrees of development, and a spiritual-divine self or monad. Evolution involves the unfolding of latent powers and capacities in response to impulses from within and stimuli from without, and the development of suitable physical forms through which they can be expressed.
Theosophy rejects the Darwinian theory of common descent, i.e. the notion that the first unicellular organisms have undergone successive bodily transformations, leading eventually to all the present creatures on earth, including humans. The enormous gaps in the fossil record prove that there has been no continuous, gradual transformation of one species into a higher species. Darwinists of the ‘punctuationist’ school recognize this and argue that new species arise relatively suddenly in small, isolated populations, and that such episodes of speciation are separated by long periods of stasis, in which species undergo little change. However, their insistence that these sudden spurts of evolutionary creativity are ultimately based on blind chance places great strains on our credulity.
Guided and directed macromutations could result in evolutionary jumps and allow a member of one species to give birth to a more advanced species, but not if the mutations were limited to genes, which code for the synthesis of proteins but do not determine bodily form – something attributed to as yet ill-defined ‘epigenetic’ (nongenetic) factors. Moreover, this would not mean that one great stock of animals evolves into another in the Darwinian sense. For instance, a species of reptile did not give birth to the first mammal. As H.P. Blavatsky says, ‘the admitted chasm between the systems of reproduction of the oviparous vertebrates and mammalia, constitutes a hopeless crux to those who, with the evolutionists, seek to link all existing organic forms in a continuous line of descent’. She adds:
The ‘Unity of Type’ common, in a sense, to all the animal and human kingdoms, is not ... a proof of the consanguinity of all organic forms, but a witness to the essential unity of the ‘ground plan’ Nature has followed in fashioning her creatures.1
New types of creatures ‘evolve’ first on the ethereal or astral plane, based on past evolutionary forms stored in the memory of nature. Instead of an evolutionary process ‘guided’ by blind chance, as in the Darwinist scheme, and instead of new kinds of creatures being created out of nothing by a single supreme being or ‘God’, as in the creationist scheme, theosophy postulates an all-pervasive instinctive intelligence, arising from the workings of a hierarchy of agencies, from higher intelligences to elemental nature-forces, including soul-impulses from within each organism, reflecting its own evolutionary needs.
Theosophy says that the present earth is the reembodiment of a former earth, and the different classes or kingdoms of monads now evolving on it and forming it – from submineral (elemental) to superhuman – are pursuing an evolutionary journey that has no absolute beginning and will have no absolute end. The present earth and its life forms originated in a highly ethereal condition and gradually materialized and condensed during the ‘descending arc’ of the earth’s evolution, which lasted until the midpoint of the earth’s lifespan, some 4.5 million years ago, in the middle of the current, fourth round of evolutionary activity. Since then the ascending arc of etherealization and spiritualization has begun.
During each of the seven rounds, the various kingdoms of monads, or life-waves, pass successively through the 12 globes of the earth’s planetary chain; our own globe is the lowest and densest, while the others are located on higher, more ethereal planes. The first root-race, or humanity, in the current round began to develop in the mid-Palaeozoic; these early protohuman forms were huge, ovoid, semi-astral, nonselfconscious beings, which slowly materialized, declined in size, and assumed the present human shape during the ensuring millions of years.
In the late second and early third root-races, reproduction took place by budding or gemmation; vital cells were thrown off from the ethereal bodies of the time and developed either into other humans or into the beginnings of the mammals, according to their inherent evolutionary tendencies. This accounts for the general resemblance between their embryonic stages and those of man. Sexual reproduction in the human kingdom is said to have originated in the second half of the third, Lemurian root-race, some 18½ million years ago. On the theosophical timescale, this was in the late Jurassic of the Mesozoic era, or age of reptiles. By this time the outer human body was becoming distinctly physical, and also sufficiently developed for the emergence of self-consciousness to begin.2
H.P. Blavatsky and G. de Purucker cited various pieces of evidence contradicting the ape-ancestry theory, including finds of human remains, footprints and stone tools, such as those presented in section 3, showing that humans of one type or another existed in much earlier geologic periods than orthodox evolutionary theory allows,1 and evidence such as that mentioned in the previous section, showing that the modern human anatomy is simpler and less specialized than that of our supposed ancestors.2 They also mentioned several contemporary scientists who believed that apes had evolved from man, while man had evolved from other types of animals.3 Blavatsky called the ape-ancestry hypothesis ‘self-degrading’ and ‘the most extravagant theory of the ages’.4
Theosophy teaches that the lower simians, the monkeys, arose from interbreeding between a ‘mindless’ or unselfconscious stock of the third root-race (who were apelike in appearance*) and a high animal stock, while the anthropoid apes resulted from interbreeding between a less progressed stock among the fourth, Atlantean, root-race and the descendants of the earlier interbreeding. ‘Thus,’ says G. de Purucker, ‘the apes and the monkeys have traces of human blood in their veins; the monkeys a single dose, so to say, of the nobler strain, and the apes a double dose of the same. But no man has one drop of either simian or anthropoid blood in his veins.’5
*Human embryos develop by the fourth week a well-defined tail. It reaches its maximum length in the sixth week when it may have as many as 12 vertebrae, after which it begins to regress, becoming the coccyx (tailbone). The coccyx in humans and apes therefore represents a vestigial tail. However, this does not require a belief in a simian ancestry, for theosophy says that the ethereal human forms in the third round were apelike and had a short tail (the sixth week of embryonic development also corresponds to the mid-second and early third root-races of the present round). The loss of a tail has actually reached a more advanced stage in the gorilla than in man, because the gorilla’s coccyx consists of only three caudal vertebrae whereas in humans it generally consists of four or five.6
The ‘humans’ involved in producing the ancestors of the monkeys were devoid of self-consciousness and acted instinctually, and hence the interbreeding was not a crime. The animals involved, like all mammals, had originally sprung from ‘man’, and the interbreeding led to fertile offspring because the distance between the humans and animals concerned was not yet very great. The Atlanteans who later repeated the act were aware of what they were doing and therefore committed deliberate bestiality. Even then, these humans and animals were not far enough apart to prevent fertile mating.
In far past geological times both these simian stocks resembled their respective human half-parents much more closely than do their present-day descendants, the living monkeys and apes. The earlier stocks were far nearer in time to the dominant human influence in their heredity, while the living simians show the effects of specialization away from that influence over the intervening millions of years.
Referring to the Lemurian episode of interbreeding, the Stanzas of Dzyan7 state that ‘those who had no spark’ (i.e. nonselfconscious humans) ‘took huge she-animals unto them’,* which belonged to several species and were quite different from any known today, and ‘begat upon them dumb races’. The offspring are described as a ‘race of crooked, red-hair-covered monsters, going on all fours’, ‘a truly pithecoid species, now extinct’, and a Commentary mentions: ‘Red-haired, swarthy men going on all-fours, who bend and unbend (stand erect and fall on their hands again) who speak as their forefathers, and run on their hands as their giant fore-mothers.’8
*Although reference is usually made to male Lemurians or Atlanteans mating with female animals,9 it’s hard to believe that no human females ever succumbed to the same vice!
Another stanza says that early Lemuro-Atlanteans also mated with a lower race of still mindless humans, who were very dissimilar physically and mentally to the more developed races. The semi-humans, or ‘so-called animals’, are described by a Commentary as a biped of human shape, but covered with hair below the waist.10 We are also told that the egg-like shells in which humans gestated during the early third root-race were often tampered with by huge animals resulting in semi-human monsters, and that certain Lemurians later interbred with some of these giant creatures.11
As regards the period in which the events in question took place, Blavatsky says that occultism ‘traces some of the most anthropoid species to the third race man of the early Atlantean period’. The later third race overlaps with the early Atlantean race, and the events could have taken place in the Cretaceous, after most humans had already acquired self-conscious minds, though De Purucker says they might have happened in the preceding Jurassic period.12 (The Cretaceous began about 16 million years ago according to theosophy, and 144 million years ago according to science.)
We are told that the descendants of the huge humanlike monsters produced by certain Lemurians ‘were modified by external conditions, until the breed, dwindling in size, culminated in the lower apes of the Miocene period. With these the later Atlanteans renewed the sin of the “Mindless” – this time with full responsibility. The resultants of their crime were the species of apes now known as Anthropoid.’13 Mating has presumably taken place with various animal or semi-animal stocks over the course of time, as the end result was not only the present monkeys and apes but also semi-human tribes which have now largely died out.14
The original apes were reportedly looked upon askance by the more evolved humans of that time but were tolerated because they had glimmerings of our active minds. They were virtually thinking entities of a lower type, and had distinct languages of their own.
The most human of these apes died out, partly because the Atlanteans, realizing the sin of their own less evolved men, made vigorous war upon them, wars of extermination; and also because the milieu, the surroundings, were not conducive to the continuance of this partly-human partly-animal race. Only the least progressed of the apes were allowed to live by the Atlanteans; and the apes today are the descendants of those who were allowed to continue to live.15
In the epic Hindu poem the Ramayana, the apes are depicted as far more humanlike than they are today; they even talk and have their own governments and laws. Led by Hanuman, the monkey god, they fight on the side of Rama against the Rakshasas of Lanka in the great epic war of India. Viewed theosophically, the depiction of the apes in the Hindu legend is not entirely fictional.
The souls that incarnated in the original monkeys and apes had not quite reached the human stage. We are told that the apes are destined to become humans of a low grade in the last two root-races of the present round and in the next or fifth round. This does not mean that ape bodies will metamorphose into human bodies, but that the souls currently inhabiting ape bodies will take on bodies among the lowest human stocks.
The earliest humans, or rather protohumans, in the present round were semi-astral beings of titanic size, and have gradually diminished in stature ever since. The Lemurians were around 7.5 m (25 ft) tall at the time of Lemuria’s destruction, as were the Atlanteans in their heyday. By about half a million years ago, man’s height had declined to about 3.5 m, and has continued to decline to about half that size today.1
The late second and early third races could conceivably have left fossils, and this is even more true of the more solidified bodies of the later third race and the fourth race.2 However, the fossil record is extremely fragmentary; for instance, it is estimated that it provides us with evidence for only about 3% of all the primate species that have ever existed.3 There are various reasons for this. First, fossilization can occur only under exceptional conditions. Second, erosion and geological cataclysms have destroyed 90 to 99% of the fossil record. Third, many fossils are buried in inaccessible places, including deep below the present ocean floors (the Lemurian continental system was centred in the Pacific Ocean, and the Atlantean continental system in the Atlantic Ocean4). Finally, cremation is said to have been widespread among humans until about 100,000 years ago.
As already shown, many remains of giant humans have in fact been found. None are displayed in any museums, and we can only speculate on what might be stored in museum basements. Given that the prevailing orthodoxy affects how scientists perceive the world, there is no guarantee that fossil fragments of giant humans would be recognized as such; after all, ‘everyone knows’ that races of human giants exist only in fairytales! In the past, bones of large extinct animals were sometimes mistaken for remains of giant humans – but the opposite could also occur.
The first mammals appear in the fossil record in the early Mesozoic, but most of the known mammals of that era were not much larger than mice. None of the fossil mammals from the later Mesozoic seem to be suitable candidates for the huge creatures with which certain Lemurians interbred or for their offspring, the ancestors of the present monkeys. Fossils of primitive monkeys and anthropoid apes first appear in the Oligocene epoch. Oligocene deposits at Fayum, Egypt, for example, contain a variety of small fossil apes, such as Aeolopithecus, Aegyptopithecus and Propliopithecus (which some scientists speculate was one of our distant ancestors). Aegyptopithecus evolved to become about the size of a large house cat.
The Miocene saw the abrupt appearance of an incredible variety of tailless apes; about 100 species roamed Europe, Africa and Asia during that epoch, classified into about 27 genera. Nearly all the species went extinct, leaving only six to thrive: two types of gorilla, two types of chimp, and two types of orangutan. Today’s great apes are therefore ‘members of a group that’s been steadily declining since the later Miocene’.5
The size of the known Miocene apes ranged from 0.6 m (2 ft) tall to around 3.1 m (10 ft) or more in the case of Gigantopithecus. Not much is known about the bodies of Miocene apes; most of the categories have been classified solely by skulls, skull parts and teeth. Quadrupeds have arms that are distinctly longer than their legs while humans have arms that are distinctly shorter than their legs. Although some Miocene apes have arms equal in length to their legs, most scientists automatically assume that every Miocene ape was a quadruped. However, it is quite possible that at least some of them, such as Gigantopithecus and Oreopithecus, were bipedal or semi-bipedal, and that some of the wildmen from around the world are their descendants.6
As in the case of the Lemurian interbreeding, no details are given in theosophical literature about the sizes of the humans and animal species involved in the later Atlantean interbreeding and of their semi-human, semi-animal offspring. Some of the known fossil apes from the Miocene could certainly have been involved, but some simian species are said to be the product of the ordinary evolutionary process.7
The seeds of our fifth (Aryan or Indo-European) root-race began to emerge in earliest Tertiary times, 7 or 8 million years ago by theosophical reckoning. The fifth race’s satya-yuga, or golden age of innocence, began about 4.5 million years ago, in the Oligocene, when the fourth race was finishing its kali-yuga, or dark age. We are not told how tall the earliest members of our humanity were, but if some of the fossil evidence documented by Cremo and Thompson is valid, humans of approximately our own size could have existed in the Pliocene, Miocene, and even earlier; they would therefore have been pygmies compared to most of the Atlanteans of the day. The fifth race became a distinct racial stock in Central Asia in the late Pliocene, about 1 million years ago.8
Since the early Miocene many apelike and semi-human species have appeared and disappeared, and many civilizations have come and gone. Mainstream anthropology ignores much of the ‘anomalous’ evidence already presented regarding the antiquity of humanity, the existence of giants, and the survival of bipedal hominoids in remote areas of the world until the present day. The fossils of primitive humans and bipedal apes that have been discovered were not our own direct ancestors but dehumanized offshoots from the main stream of human evolution, some of them being products of various episodes of human-animal crossbreeding. They could have existed alongside advanced civilizations, just as hunter-gatherer societies coexist with more advanced cultures today.
Of all the creatures on earth, only humans are known to have self-conscious minds – the ability to remember the past and plan for the future, and to express their thoughts in a variety of creative ways. The human brain, with its 86 billion neurons and over 100 trillion synaptic connections, is the most complex arrangement of matter known. According to mainstream science, brain activity somehow gives rise to self-awareness and the power of thought. But materialists are forced to admit that they ‘have as yet no idea whatever about how a mass of electrical and chemical discharges within the brain is converted into what we individually and subjectively experience as consciousness’.1 In addition to random mutations, another ‘driving force’ behind the evolution of apes into humans is claimed to be the development of drier and cooler climatic conditions – in other words, self-conscious intelligence emerged due to a change in the weather!
Some scientists have recognized that mind ‘defies explanation in terms of evolutionary theory’ and is ‘impossible to explain in purely material terms’.2 Astrophysicist Freeman Dyson has said:
The prevailing view among biologists seems to be that the mind arose accidentally out of molecules of DNA or something. I find that very unlikely. It seems more reasonable to think that mind was a primary part of nature from the beginning and we are simply manifestations of it at the present stage of history. It’s not so much that mind has a life of its own, as that the mind is inherent in the way the universe is built, and life is nature’s way to give mind opportunities it wouldn’t otherwise have.3
Neuroscientist and Nobel Prize winner John Eccles said that the materialistic theory of mind is ‘impoverished and empty’ and fails to account for ‘the wonder and mystery of the human self with its spiritual values, with its creativity, and with its uniqueness for each of us’.4 Eccles believed that there is a mental world separate from the physical world, that the mind and brain interact, and that ‘each Soul is a new Divine creation which is implanted into the growing foetus at some time between conception and birth’.5
Theosophy, too, recognizes that ‘it requires more than a mere interplay between certain material aggregates and their environment, to call to life a fully conscious man’.6 It assigns humans a spiritual ancestry, but rejects the belief that they were created by a supernatural, extracosmic, anthropomorphic God. If nature is infinite, divinity cannot be outside nature but must be all-nature itself; and nature is synonymous with boundless consciousness-life-substance, manifesting in infinitely diverse forms. The spiritual monad at the heart of every entity embodies in an endless variety of forms in an endless variety of worlds in the course of its cyclic evolutionary development. The earth is merely the latest station on its unending evolutionary journey.
The earliest ethereal human races in the present round did not have self-conscious minds. After their outer forms had become more physical and attained the necessary degree of complexity and refinement, the gradual awakening and unfoldment of our latent intellectual and spiritual powers could begin, under the influence of a higher part of our constitution; this process is referred to allegorically as the incarnation of the manasaputras (a Sanskrit word meaning ‘sons of mind’).7
This teaching is echoed in many myths and religious allegories. In the story of Prometheus, for instance, Prometheus steals the spiritual fire of the gods (intelligence) and brings it to mankind. In punishment he is chained to a rock (our human bodies) on a mountainside, where he will remain throughout the ages until mankind rises to his level and liberates him.
In the Book of Genesis, the Garden of Eden represents the innocent, mindless, childlike state of infant humanity. The eating of the fruit from the tree of knowledge of good and evil signifies the awakening of self-consciousness, when man becomes ‘as the gods’, endowed with free will. The serpent which tempts Eve is a symbol of wisdom – hence the injunction to be ‘wise as serpents’ (Matthew 10:16). Adam and Eve, representing early humanity, are then expelled from Paradise, and ‘fall’ from their peaceful state of blissful unselfconsciousness into one of struggle, temptation and ethical responsibility in the material world. They put on ‘coats of skin’, meaning that it was in the same period as the arousing of our minds that the semi-astral bodies of early humanity became densely physical.
The awakening of mind in the present round began during the fifth subrace of the third root-race, around 18½ million years ago. However, not all humans began to gain self-consciousness at the same time or at the same pace, as they were not all at exactly the same stage of development. Even now our mental powers are still developing and will reach their full efflorescence in the following round, millions of years hence.
In a sense, theosophy agrees with science that humans and apes had a common ancestor – but that common ancestor was man himself, in his earlier, more primitive form. In the remote past – at the end of the third round and at the end of the third root-race of the present round – the human body was apelike in appearance. However, it did not evolve from animal bodies but from astral prototypes. Humans could be called ‘fallen angels’ rather than risen apes. Our spiritual monad, the ‘angel’ in us, does not literally descend into matter; it remains on its own plane and works through a series of lower vehicles. When the densest of them, the physical body, was ready, man’s latent self-conscious intelligence began to be unfolded and expressed. The physical and spiritual lines of evolution meet in the third stream of evolution – the mental. Humanity therefore forms a separate kingdom, poised midway between the animals and the superhuman kingdoms or ‘gods’.
The ape-ancestry myth: contents